Environmental Health Perspectives Volume 103, Number 1, January 1995
![]()
Environmental
Health Perspectives Volume 103, Number 1, January 1995
Department of Emergency Medicine, East Carolina University School of Medicine, Greenville, NC 27858 USA and New York City Poison Center, New York, NY 10016 USA
http://ehpnet1.niehs.nih.gov/docs/1995/103-1/meggs.html
Address correspondence to W.J. Meggs, Room 4W54, Brody Building, Department of Emergency Medicine, East Carolina University, Greenville, NC 27858 USA.Received 1 July 1994; accepted 14 September 1994.
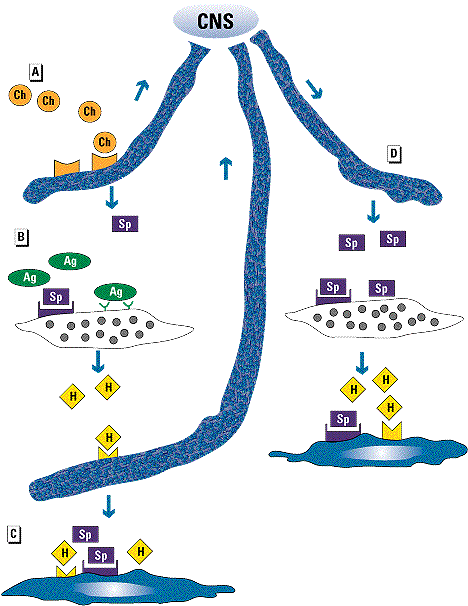
Figure 1. Interplay between neurogenic and immunogenic inflammation, with neurogenic switching of the inflammation site. (A) Chemical irritants (Ch) bind sensory neurons to release neuropeptides including substance P (Sp). An afferent signal travels to the central nervous system (CNS). (B) Antigen (Ag) binds IgE molecules on mast cells to release mast cell mediators including histamine (H). Histamine binds to receptors on sensory neurons, and substance P binds to receptors on mast cells. (C) Both histamine and substance P can bind effector cells, such as endothelial cells, mucus-secreting cells, and bronchial smooth muscle cells to produce inflammation. (D) Neurogenic switching occurs when an efferent signal from the CNS causes release of neuropeptides at another site, producing inflammation at the second site without local stimulation.One puzzling feature of the inflammatory response is that a stimulus in one tissue can sometimes lead to inflammation at another site. Food allergy provides an example. Ingestion of a food allergen can produce gastrointestinal symptoms, with diarrhea, abdominal pain, bloating, and emesis arising from the direct degranulation of gut mast cells with local mediator release. A small percentage of patients with food allergy develop symptoms at other sites, manifesting as asthma (5), rhinitis (6), or urticaria (7). Food hypersensitivity can also manifest as arthritis (8,9) and migraine (10,11). Histamine from gut mast cells could bind to sensory nerves to produce an afferent signal, which could be rerouted via the central nervous system to another site, as depicted in Figure 1. This neurogenic switching could then explain the diverse manifestations of food allergy.
Systemic anaphylaxis may be a manifestation of neurogenic switching. Cutaneous inoculation with an antigen, such as a bee sting, or gut inoculation, as in the ingestion of a food or drug, can affect multiple organ systems immediately. Respiratory involvement with bronchospasm, bronchorrhea, and laryngeal edema, gastrointestinal symptoms, skin involvement away from the site of inoculation with diffuse flushing or urticaria, and cardiovascular symptoms with hypotension from vasodilation can all arise. A role for the nervous system in systemic anaphylaxis has been demonstrated in experimental models. It is known that vagotomy protects rats from lethal anaphylaxis without changing the production of antibody or histamine release (12). Experimental lesions of the anterior hypothalamus lessens the anaphylactic reaction in a guinea pig model (13). Neurogenic switching may be the mechanism for this observed modulation of anaphylaxis by the nervous system, though other mechanisms such as a generalized decrease in parasympathetic tone should be considered.
Gustatory rhinitis is another phenomenon in which neurogenic switching may play a role. In this syndrome, rhinorrhea, nasal congestion, and facial sweating develop after the ingestion of spicy foods. Ingested irritants such as capsaicin, the active ingredient in chili peppers, interact with branches of the trigeminal nerve innervating the oral cavity. The efferent signal is switched to the nose and face (14).
Sick building syndrome (15), in which diverse symptoms such as headache and difficulty with concentration accompany mucosal irritation in occupants of poorly ventilated buildings, may involve neurogenic switching. The site of inflammation could be switched from the airway to the brain, causing vasodilation and edema. An alternative hypothesis is that lymphocytes at the site of stimulation release mediators which circulate to the brain and cause symptoms (16). Interleukin-1 and interleukin-2 are known to affect cerebral function in animal models (17,18). This type of switching might be termed "immunogenic switching."
Neurogenic switching may play a role in multiple chemical sensitivity syndrome (19-21), which is thought to be mediated by neurogenic inflammation (22). In this syndrome, exposure to respiratory irritants triggers symptoms in more than one organ system. An individual patient with chemical sensitivities often has recurrent sites of symptomatology and inflammation which reoccur with a well-defined pattern. Myalgias in the nape of the neck, inflammation of a particular set of joints, or gastrointestinal symptoms might occur over and over again in a single patient. The establishment of a neuronal pathway so that stimulation of irritant receptors in the airway, for example, leads to inflammation in a given tissue may be the mechanism for involvement of secondary organ systems.
Neurogenic inflammation is thought to play a role in rheumatoid arthritis (23), migraine headache (24,25), and fibromyalgia (26). The basis of these conditions may be an acquired neuronal pathway that shunts inflammatory stimuli to the joints, cerebral vasculature, or muscles, respectively. Then, inflammatory stimuli arising from allergens, chemical irritants, or infectious agents could lead to a flare in inflammation at the diseased site. Emotional stress might also lead to neuronal signals that could result in inflammation at susceptible sites.
To summarize, it has been pointed out that the multiple organ system involvement that has been so problematic in understanding chemical sensitivity also occurs in allergy. A single mechanism may underlie this switching of the site of inflammation in both allergy and chemical sensitivity. It is hypothesized that neurogenic switching is one possible mechanism by which stimulation of inflammation at one site can lead to inflammation at another (Fig. 1). An exposure to either an allergen or chemical irritant at one site leads to a sensory nerve impulse. For allergens, mast cell degranulation leads to the release of histamine and other mediators, and histamine binds receptors on sensory nerves. For chemical irritants, receptors on peripheral nerves are directly triggered. When the impulse reaches the central nervous system, it is redirected to another peripheral location, leading to the release of substance P and other neuropeptides, producing inflammation at the second site.
An alternative hypothesis is that immunogenic switching occurs, with the release of cytokines which act on distant cells. There are several differences expected between neurogenic switching and immunogenic switching. The time required for the onset of neurogenic switching depends on nerve conduction velocity, while immunogenic switching depends on circulation time and diffusion times in tissues. Neurogenic switching would directly target a particular organ in a repetitive pattern, while immunogenic switching might have a more diffuse effect. The mechanism by which the site of inflammation is shifted in both allergy and chemical sensitivity can be determined experimentally, both in controlled challenges in human subjects and experimental models. Such a program could further our understanding of these conditions while leading to improvements in diagnosis and treatment.
References
1. Foreman JC. Peptides and neurogenic inflammation. Br Med Bull 43:386 (1987).
2. Hinsey JC, Gasser HS. The component of the dorsal root mediating vasodilation and the Sherrington contracture. Am J Physiol 92:679 (1930).
3. Chahl LA. Antidromic vasodilation and neurogenic inflammation. Pharmacol Ther 37: 275-300 (1988).
4. Cavagnaro J, Lewis RM. Bidirectional regulatory circuit between the immune and neuroendocrine systems. Year Immunol 4:241-252 (1989).
5. Onorato J, Merland N, Terral C, Michel FB, Bousquet J. Placebo-controlled double-blind food challenge in asthma. J Allergy Clin Immunol 78:1139-1146 (1986).
6. Bousquet J, Chanez P, Michel F-B. The respiratory tract and food hypersensitivity. In: Food allergy: adverse reactions to foods and food additives (Metcalfe DD, Sampson HA, Simon RA, eds). Oxford:Blackwell Scientific Publications,1991;139-149.
7. Adkins FM. Food-induced urticaria. In: Food allergy: adverse reactions to foods and food additives (Metcalfe DD, Sampson HA, Simon RA, eds). Oxford:Blackwell Scientific Publications,1991;129-138.
8. Parke AL, Hughes GRV. Rheumatoid arthritis and food: a case study. Br Med J 282: 2027-2029 (1981).
9. Panush RS, Stroud RM, Webster EM. Food induced (allergic) arthritis: clinical and serological studies. Arthritis Rheum 29:220 (1986).
10. Egger J, Wilson J, Carter CM, Turner MW. Is migraine food allergy? A double-blind trial of oligoantigenic diet treatment. Lancet 2: 865-869 (1983).
11. Mansfield LE, Vaughan TR, Waller SF, Haverly RW, Ting S. Food allergy and adult migraine: double blind and mediator confirmation of an allergic etiology. Ann Allergy 55: 126-129 (1985).
12. Levy RM, Rose JE, Johnson JS. Effect of vagotomy on anaphylaxis in rat. Clin Exp Immunol 24:96-101 (1976).
13. Leslie CA, Mathe AA. Modification of guinea pig lung anaphylaxis by central nervous system (CNS) perturbations. J Allergy Clin Immunol 83:94-101(1989).
14. Raphael GD, Baraniuk JN, Kaliner MA. How and why the nose runs. J Allergy Clin Immunol 87:457-467 (1991).
15. Molhave L. The sick buildings--a subpopulation among the problem buildings. In: Indoor Air '87. Proceedings of the IV International conference on indoor air quality and climate, vol 2 (Sefert B, Esdorn H, Fischer M, eds). Berlin:Institute for Water, Soil, and Air Hygiene,1987.
16. Meggs WJ. Multiple chemical sensitivities and the immune system. Toxicol Ind Health 8: 203-214 (1992).
17. Obal F Jr, Opp M, Cady AB. Interleukin 1 alpha and interleukin 1 beta fragments are somnogenic. Am J Physiol 259:R439-446 (1990).
18. DeSarro GB, Masuda Y, Ascioti C, Audino MG, Nistico G. Behavioral and ECG spectrum changes induced by intracerebral infusion of interferons and interleukin-2 in rats are antagonized by naloxone. Neuropharmacology 29: 167-179 (1990).
19. Randolph TG. Human ecology and susceptibility to the chemical environment. Springfield, IL:C. C. Thomas, 1962.
20. Tabershaw IR, Cooper WC. Sequelae of acute organic phosphate poisoning. J Occup Med 8:5-19 (1966).
21. Cullen MR. The worker with chemical sensitivities: an overview. State Art Rev Occup Med 2:655-661 (1987).
22. Meggs WJ. Neurogenic inflammation and sensitivity to environmental chemicals. Environ Health Perspect 101:234-238 (1993).
23. Gronblad M, Konttinen YT, Korkala O, Liesi P, Hukkanen M, Polak JM. Neuropeptides in synovium of patients with rheumatoid arthritis and osteoarthritis. J Rheum 15:1807-1810 (1988).
24. Silberstein SD. Advances in understanding the pathophysiology of headache. Neurology 42(suppl 2):6-10 (1992).
25. Buzzi MG, Moskowitz MA. The trigemino-vascular system and migraine. Pathol Biol 40:313-317 (1992).
26. Zimmerman M. Pathophysiologic mechanisms of fibromyalgia. Clin J Pain 7(suppl):S8-15 (1991).
Last Update: May 21, 1998