Toxicological
Defense Mechanisms and the Shape of Dose-Response Relationships
Environmental
Health Perspectives 106, Supplement 1, February 1998
Center for Environmental Health and Occupational Medicine, Department of Pharmacology, Toxicology, and Therapeutics, University of Kansas Medical Center, Kansas City, Kansas
Key words: cadmium, carbon tetrachloride, tolerance, hepatotoxicity, metallothionein induction, subcellular distribution, sequestration, MT-transgenic animals, MT-null mice
This paper is based on a presentation at The Third BELLE Conference on Toxicological Defense Mechanisms and the Shape of Dose-Response Relationships held 12-14 November 1996 in Research Triangle Park, NC. Manuscript received at EHP 7 March 1997; accepted 17 July 1997.
This work was supported by National Institutes of Health grants ES-01142 and ES-07079. We thank the following graduate and postdoctoral fellows who contributed to this project: S.T. Cagen, P. Goering, S. Habeebu, M.B. Iszard, and Y.P. Liu. We also thank R.D. Palmiter, K.H.A. Choo, and G.K. Andrews for their collaboration in these studies.
Address correspondence to Dr. C.D. Klaassen, Department of Pharmacology, Toxicology, and Therapeutics, University of Kansas Medical Center, Kansas City, KS 66160-7417. Telephone: (913) 588-7714. Fax: (913) 588-7501. E-mail: cklaasse@kumc.edu
Abbreviations used: CCl4, carbon tetrachloride; MT, metallothionen.
MT-I and -II can be induced easily by heavy metals, hormones, inflammation, acute stress, and many chemicals (1). In essence induction of MT has been proposed as an important adaptive mechanism in response to environmental stimuli. Induction of MT protects against metal toxicity (6), acts as a free radical scavenger protecting against oxidative damage (7), and protects against toxicity of alkylating anticancer drugs and other electrophiles (8).
In this paper we demonstrate that induction of MT is an important cellular adaptive mechanism protecting against the toxicity produced by metals such as Cd as well as by organic chemicals such as carbon tetrachloride (CCl4).
There have been several hypotheses proposed to explain Cd-induced tolerance. Originally it was hypothesized that Cd pretreatment alters the organ distribution of Cd, with more Cd distributing to the liver and less to the kidney. However no major differences in the distribution of Cd to various organs have been observed between control and Cd-pretreated animals (12). It was also hypothesized that tolerance to Cd-induced lethality is attributable to increased biliary excretion of Cd, which has not turned out to be true either. Pretreatment of animals with Cd or Zn actually decreased or prevented the biliary excretion of Cd (14,15).
Why do animals die from acute Cd toxicity? It was originally thought that animals died from Cd-induced cardiotoxicity or nephrotoxicity. However we know now that this is not true. In fact we showed that animals exposed to acute high doses of Cd probably die from liver injury (16). The liver accumulates substantial amounts of Cd after both acute and chronic exposure (6,17). Cd produces dose-dependent liver injury in laboratory animals within 10 hr after iv administration (16-19), with congestion, apoptosis, necrosis, and peliosis as major features of injury (16-20). The Cd-induced liver injury is so severe that hepatic failure is believed to be responsible for acute Cd lethality (12,16,18).
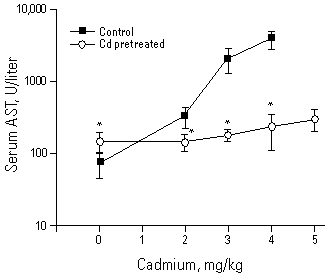
Tolerance to acute Cd toxicity is apparently attributable to MT protection against Cd-induced acute hepatotoxicity (10,12,13). Indeed, after treatment of animals with a low dose of Cd, the liver injury caused by a subsequent toxic dose of Cd is markedly reduced (Figure 1). The hepatoprotection is not attributable to a decreased accumulation of Cd in the liver (12). However, the subcellular distribution of Cd is dramatically altered, with more Cd distributing to the cytosol and a significant reduction of Cd in critical organelles such as nucleus, mitochondria, and microsomes (Figure 2). Chromatography of the cytosolic fraction indicates that most of the Cd in Cd-pretreated animals is associated with MT (Figure 3). Thus the protective role of MT in Cd tolerance to hepatic injury is proposed.
Figure 1. The protective effect of CdCl2 pretreatment (2.0 mg/kg Cd sc for 24 hr) against the hepatotoxic effects of the subsequent high doses of Cd challenge (2.0-5.0 mg/kg Cd iv for 10 hr). Liver injury was measured by plasma aspartate AST. The 5.0 mg/kg dose was not used in control rats as preliminary studies indicate a high rate of mortality in this group. Values represent mean±SE of 4 to 6 rats. *Significantly different from controls at p<0.05. Abbreviation: AST, aminotransferase activity. Reproduced from Goering and Klaassen (12); with permission of Academic Press.

Figure 2. Hepatic subcellular distribution of 109CdCl2 2 hr after challenge (3.5 mg/kg Cd iv) following saline or Cd (2.0 mg/kg sc for 24 hr) pretreatment. The cellular pellets were defined as Nuc (600g, 10 min), Mit (10,000

Figure 3. Representative gel-filtration elution profiles of 109CdCl2 in the hepatic cytosols 2 hr after challenge (3.5 mg/kg Cd iv) in control or Cd (2.0 mg/kg sc for 24 hr) pretreated rats. Radioactive Cd eluting with retention coefficients (Ve/Vo) of 1.0 to 1.5 and 1.75 to 2.25 are Cd bound to high-molecular-weight proteins and MT, respectively. Reproduced from Goering and Klaassen (11), with permission of Academic Press.
Newborn animals have high concentrations of MT in their livers; thus they are resistant to Cd-induced lethality and hepatotoxicity. For example Cd treatment (4.0 mg/kg) produced a 20-fold increase in serum alanine aminotransferase activity in adult rats, but in newborns (10-day-old rats) 6 mg Cd/kg did not produce liver injury (21).
Recently we demonstrated that MT-I transgenic mice, which have a 10-fold higher concentration of MT in their liver than control mice (22), are resistant to Cd-induced lethality and hepatotoxicity, as evidenced by >90% lower activities of serum alanine aminotransferase and sorbitol dehydrogenase (Figure 4). In contrast MT-null mice have an increased susceptibility to Cd-induced lethality and hepatotoxicity (23-25), and liver injury is more severe in MT-null mice than in corresponding controls (Figure 5). Furthermore Zn pretreatment, which increases hepatic MT 20-fold in control but not in MT-null mice, protects against Cd-induced hepatotoxicity in control but not in MT-null mice (Figure 5), thus supporting our earlier observation that Zn-induced tolerance to Cd is attributable to induction of MT (26).
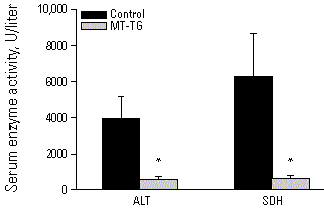
Figure 4. Serum ALT and SDH activities in control and MT-TG mice 24 hr after injection of a hepatotoxic dose of CdCl2 (3.1 mg/kg Cd iv). Values represent the mean ± SE of 16 to 25 mice. *Significantly different from controls at p<0.05. Abbreviations: ALT, alanine aminotransferase; SDH, sorbitol dehydrogenase; MT-TG, MT-transgenic mouse. Reproduced from Liu et al. (22), with permission of Academic Press.
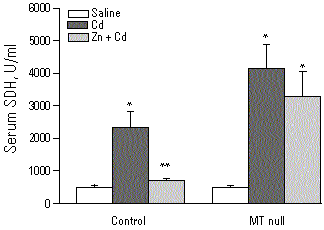
Figure 5. SDH activities in control and MT-null mice pretreated with saline or Zn (200 µmol/kg, sc
These data indicate that both constitutive and inducible MT are responsible for the detoxication of Cd. Pharmacodynamic tolerance occurs via high-affinity sequestration of the metal within the cell. As a result most of the Cd in cells is bound to MT in the cytosol, with a concomitant reduction of the Cd available to bind/damage critical organelles (6). Using MT-null mice, we find that intracellular MT also plays an important protective role in chronic Cd nephrotoxicity (J Liu et al., in preparation). Binding of metal ions to MT also appears to be the mechanism for the protection against the toxicity of other metals such as mercury, silver, and cisplatin (27,28). Thus, induction of MT is an important adaptive mechanism preventing metal toxicity in animals as well as in humans.
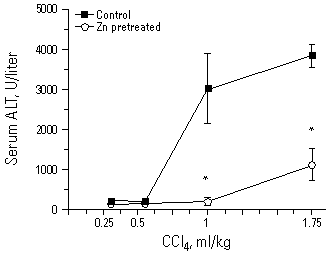
Figure 6. The protective effect of Zn pretreatment (ZnCl2 250 µmol/kg ip) against the hepatotoxicity of carbon tetrachloride (0.3-1.75 ml/kg in corn oil, ip for 24 hr). Liver injury was measured by serum ALT. Values represent mean±SE of 4 to 6 rats. *Significantly different from controls at p<0.05. Reproduced from Goering and Klaassen (30), with permission of Academic Press.
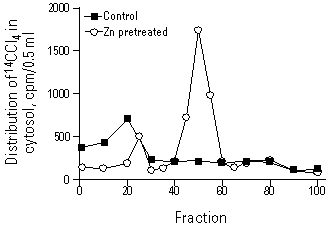
Figure 7. Representative gel-filtration elution profiles of 14C-Cl4 in the hepatic cytosols 90 min after administration of 14C-Cl4 (25 µCi/kg ip) in control or Zn (250 µmol/kg, ip for 24 hr) pretreated rats. Radioactive 14C-Cl4 eluting in fractions 15 to 25 and fractions 45 to 55 are 14C-Cl4 bound to high-molecular-weight proteins and MT, respectively. Reproduced from Goering and Klaassen (30), with permission of Academic Press.
The potential role of MT induction as an adaptive mechanism decreasing CCl4 toxicity is further supported by other studies. First, 24-hr pretreatment with Zn increased hepatic MT and protected against CCl4 toxicity, while 2-hr Zn pretreatment prior to the induction of MT synthesis failed to offer protection (31). Second, mild Zn deficiency interferes with MT synthesis and thus decreases the efficacy of MT induction by turpentine to protect against CCl4 toxicity (32). Third, recent studies showed that MT-null mice are more susceptible than controls to CCl4 hepatotoxicity (33,34), indicating that MT functions as an adaptive mechanism to decrease the toxicity of CCl4.
Evidence also suggests a role for MT in protection aginst oxidative stress. MT can serve as a sacrificial scavenger for hydroxyl radicals in vitro (35) and protect against free radical-induced DNA damage (36-38). MT can also assume the function of superoxide dismutase in yeast (39) and protect against lipid peroxidation in erythrocyte ghosts produced by xanthine oxidase-derived superoxide anion and hydrogen peroxide (40). Hepatocytes from MT-null mice are more sensitive than control cells to oxidative damage produced by t-butylhydroperoxide and paraquat (41,42). MT is induced by oxidative stress-producing chemicals (43) and thus may protect against oxidative damage (7) and the toxicity of alkylating anticancer drugs (8).
References
1. Kägi JHR. Evolution, structure and chemical activity of class I metallothioneins: An overview. In: Metallothionein III: Biological Roles and Medical Implications (Suzuki KT, Imura N, Kimura M, eds). Berlin:Birkhauser Verlag, 1993;29-56.
2. Hamer DH. Metallothionein. Annu Rev Biochem 55:913-951 (1986).
3. Searle PF, Davison BL, Stuart GW, Wilkie TM, Norstedt G, Palmiter RD. Regulation, linkage, and sequence of mouse metallothionein I and II genes. Mol Cell Biol 4:1221-1230 (1984).
4. Palmiter RD, Findley SD, Whitmore TE, Durnam DM. MT-III, a brain specific member of the metallothionein gene family. Proc Natl Acad Sci USA 89:6333-6337 (1992).
5. Quaife C, Findley SD, Erickson JC, Kelly EJ, Zambrowicz BP, Palmiter RD. Induction of a new metallothionein isoform (MT-IV) occurs during differentiation of stratified squamous epithelia. Biochemistry 33:7250-7259 (1994).
6. Goering PL, Waalkes MP, Klaassen CD. Toxicology of cadmium. In: Toxicology of Metals: Biochemical Aspects, Handbook of Experimental Pharmacology. Vol 115 (Goyer RA, Cherian MG, eds). New York:Springer-Verlag, 1995;189-213.
7. Sato M, Bremner I. Oxygen free radicals and metallothionein. Free Radic Biol Med 14:325-337 (1993).
8. Lazo JS, Pitt BR. Metallothionein and cell death by anticancer drugs. Annu Rev Pharmacol Toxicol 35:635-653 (1995).
9. Terhaar CJ, Roudabush VE, Fassett DW. Protective effects of low doses of cadmium chloride against subsequent high oral doses in the rat. Toxicol Appl Pharmacol 7:500-505 (1965).
10. Leber AP, Miya TS. A mechanism for cadmium- and zinc induced tolerance to cadmium toxicity: involvement of metallothionein. Toxicol Appl Pharmacol 37:403-414 (1976).
11. Goering PL, Klaassen CD. Altered subcellular distribution of cadmium following cadmium pretreatment: possible mechanism of tolerance to cadmium-induced lethality. Toxicol Appl Pharmacol 70:195-203 (1983).
12. Goering PL, Klaassen CD. Tolerance to cadmium-induced hepatotoxicity following cadmium pretreatment. Toxicol Appl Pharmacol 74:308-313 (1984).
13. Onosaka S, Tanaka K, Cherian MG. Effects of cadmium and zinc on tissue levels of metallothionein. Environ Health Perspect 54:67-72 (1984).
14. Cherian MG. Biliary excretion of cadmium in rat. II: The role of metallothionein in the hepatobiliary transport of cadmium. J Toxicol Environ Health 2:955-961 (1977).
15. Klaassen CD. Effect of metallothionein on hepatic disposition of metals. Am J Physiol 234:E47-E53 (1978).
16. Dudley RE, Svoboda DJ, Klaassen CD. Acute exposure to cadmium causes severe liver injury in rats. Toxicol Appl Pharmacol 65:302-313 (1982).
17. Dudley RE, Gammal LM, Klaassen CD. Cadmium-induced hepatic and renal injury in chronically exposed rats: likely role of hepatic cadmium-metallothionein in nephrotoxicity. Toxicol Appl Pharmacol 77:414-426 (1985).
18. Hoffmann EO, Cook JA, DiLuzio NR, Coover JA. The effects of acute cadmium administration in the liver and kidney of the rat: light and electron microscopic studies. Lab Invest 32:655-664 (1975).
19. Dudley RE, Svoboda DJ, Klaassen CD. Time course of cadmium-induced ultrastructural changes in rat liver. Toxicol Appl Pharmacol 76:150-160 (1984).
20. Habeebu SSM, Liu J, Klaassen CD. Cadmium-induced apoptosis in mouse liver. Fundam Appl Toxicol (Suppl) 30:856 (1996).
21. Goering PL, Klaassen CD. Resistance to cadmium-induced hepatotoxicity in immature rats. Toxicol Appl Pharmacol 74:321-329 (1984).
22. Liu YP, Liu J, Iszard MB, Andrews GK, Palmiter RD, Klaassen CD. Transgenic mice that overexpress metallothionein-I are protected from cadmium lethality and hepatotoxicity. Toxicol Appl Pharmacol 135:222-228 (1995).
23. Masters BA, Kelly EJ, Quaife CJ, Brinster RL, Palmiter RD. Targeted disruption of metallothionein I and II genes increases sensitivity to cadmium. Proc Natl Acad Sci USA 91:584-588 (1994).
24. Michalska AE, Choo KHA. Targeting and germ-line transmission of a null mutation at the metallothionein I and II loci in mouse. Proc Natl Acad Sci USA 90:8088-8092 (1993).
25. Liu J, Liu YP, Michalska AE, Choo KHA, Klaassen CD. Metallothionein plays less of a protective role in CdMT-induced nephrotoxicity than CdCl2-induced hepatotoxicity. J Pharmacol Exp Ther 276:1216-1223 (1996).
26. Goering PL, Klaassen CD. Zinc-induced tolerance to cadmium hepatotoxicity. Toxicol Appl Pharmacol 74:299-307(1984).
27. Liu J, Kershaw WC, Klaassen CD. The protective effect of metallothionein on the toxicity of various metals in rat primary hepatocyte culture. Toxicol Appl Pharmacol 107:27-34 (1991).
28. Kraker A, Schmidt J, Kerzoski S, Petering DH. Binding of cis-dichlorodiammine platinum(II) to metallothionein in Erlich cells. Biochem Biophys Res Commun 130:786-792 (1985).
29. Min KS, Terano Y, Onosaka S, Tanaka K. Induction of metallothionein synthesis by menadione or carbon tetrachloride is independent of free radical production. Toxicol Appl Pharmacol 113:74-79 (1992).
30. Cagen SZ, Klaassen CD. Protection of carbon tetrachloride-induced hepatotoxicity by zinc: role of metallothionein. Toxicol Appl Pharmacol 52:107-116 (1979).
31. Clarke IS, Lui EM. Interaction of metallothionein and carbon tetrachloride on the protective effect of zinc on hepatotoxicity. Can J Physiol Pharmacol 64:1104-1110 (1986).
32. DiSilvestro RA, Carlson GP. Effects of mild zinc deficiency, plus or minus acute phase response, on CCl4 hepatotoxicity. Free Radic Biol Med 16:57-61 (1994).
33. Sato M, Naganuma A, Miura N, Tohyama C, Homma Y. Unpublished data.
34. Liu YP, Hartley DP, Liu J. Protection against carbon tetrachloride hepatotoxicity by oleanolic acid is not mediated through metallothionein. Toxicol Lett (in press).
35. Thornalley PJ, Vasäk M. Possible role for metallothionein in protection against radiation-induced oxidative stress: kinetics and mechanism of its reaction with superoxide and hydroxyl radicals. Biochim Biophys Acta 27:36-44 (1985).
36. Abel J, Ruiter N. Inhibition of hydroxyl-radical-generated DNA degradation by metallothionein. Toxicol Lett 47:191-196 (1989).
37. Chubatsu LS, Meneghini R. Metallothionein protects DNA from oxidative damage. Biochem J 291:193-198 (1993).
38. Schwarz MA, Lazo LS, Yalowich JC, Allen WP, Whitmore M, Bergonia HA, Tzeng E, Billiar T, Robbins PD, Lancaster JR et al. Metallothionein protects against the cytotoxic and DNA-damaging effects of nitric oxide. Proc Natl Acad Sci USA 92:4452-4456 (1995).
39. Tamai KT, Gralla EB, Ellerby LM, Valentine JS, Thiele DJ. Yeast and mammalian metallothioneins functionally substitute for yeast copper-zinc superoxide dismutase. Proc Natl Acad Sci USA 90:8013-8017 (1993).
40. Thomas JP, Bachowsk GJ, Girotti AW. Inhibition of cell membrane lipid peroxidation by cadmium- and zinc-metallothioneins. Biochim Biophys Acta 884:448-461 (1986).
41. Lazo JS, Kondo Y, Dellapiazza D, Michalska AE, Choo KHA, Pitt BR. Enhanced sensitivity to oxidative stress in cultured embryonic cells from transgenic mice deficient in metallothionein I and II genes. J Biol Chem 270:5506-5510 (1995).
42. Zheng H, Liu J, Liu YP, Klaassen CD. Hepatocytes from MT-null mice are sensitive to oxidative stress induced by t-butylhydroperoxide and cadmium. Toxicol Lett 87:139-145 (1996).
43. Bauman JW, Liu J, Liu YP, Klaassen CD. Increase in metallothionein produced by chemicals that induce oxidative stress. Toxicol Appl Pharmacol 110:347-354 (1991).
Last Update: March 11, 1998